
Omar López-Ramírez
Vestibular primary afferents form calyceal endings surrounding type I hair cells (HC), bouton endings contacting type II HC, or both. In vivo recordings from vestibular afferent nerve fibers (Goldberg et al., 1990) have functional differences correlated with the epithelial zones (striola, S, and extrastriola, ES) in the utricle. We are investigating the mechanisms underlying differences in spike timing and response dynamics of afferents from S and ES zones in the utricle. Low-voltage-activated K (KLV) channels (Kv1 and Kv7) and HCN channels are present in the calyx terminals and have been implicated in vestibular afferent spike regularity (Perez et al., 2009; Kalluri et al., 2010; Lysakowski et al., 2011; Horwitz et al., 2014; Meredith & Rennie, 2015) and recent evidence implicated them in the non-quantal (NQ) transmission between hair cells to calyx (Contini et al., 2017). In calyceal afferents KLV and h current have larger amplitude in the striola; in the striola, XE991 blocks KLV and h current, without changing the regularity; In LES, Kv1 and Kv7 channels contribute to the excitability, regularity and rate (fig.1).
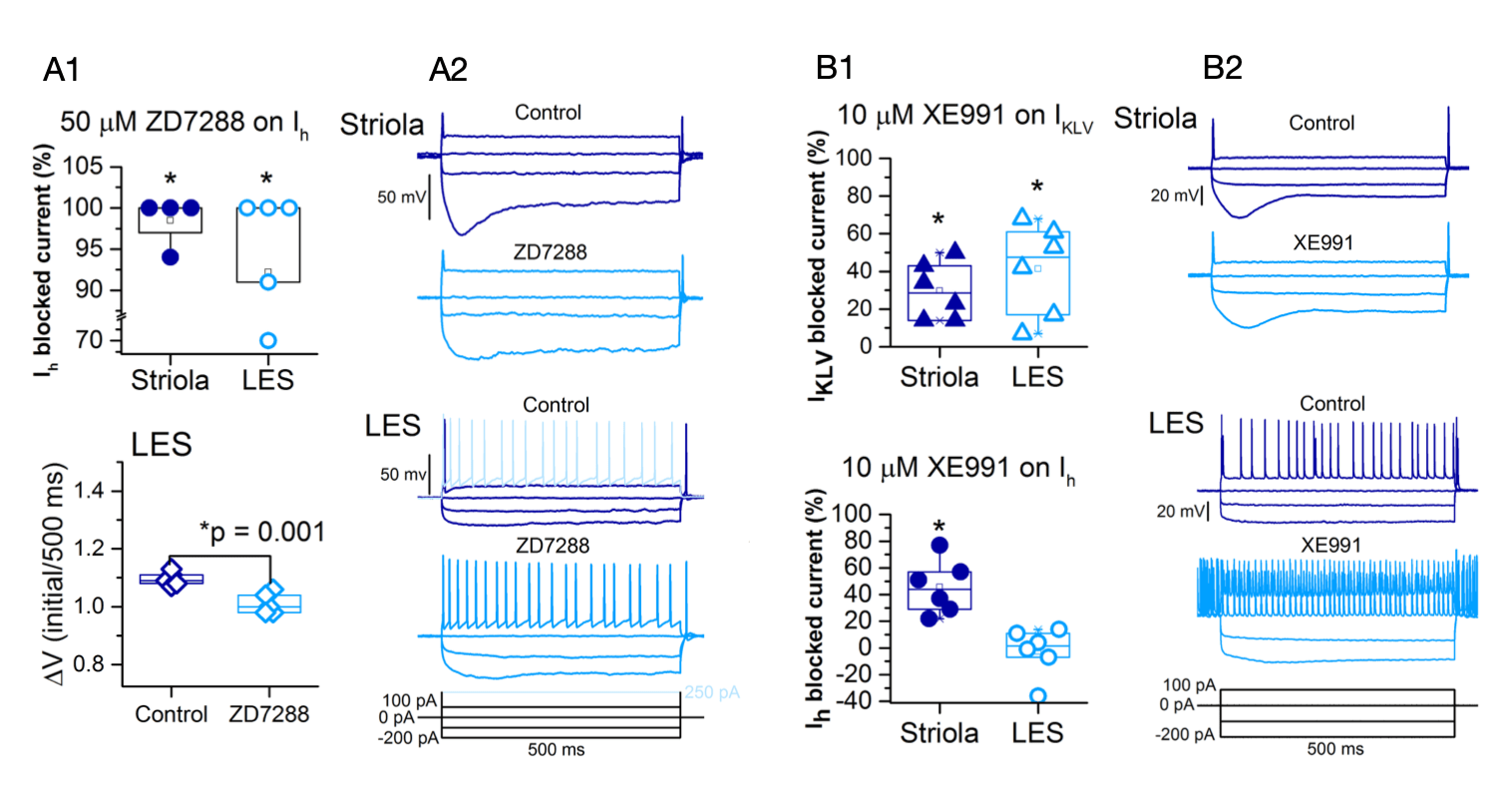
Figure 1. A1, ZD7288 significantly blocked Ih in striola (p = 0.04) and LES (p=0.03). This blockage was also evident in the voltage sag (DV), which was significantly reduced in LES (p=0.01). A2, current clamp recordings show than blocking Ih does not change the voltage threshold nor the spike regularity in neither LES or S calyces. B1, KLV current was significantly blocked by XE991 in both regions (LES p=0.0006; S p=0.02). Additionally, Ih was also significantly blocked by XE991 in striolar calyces (p=0.004). B2, current clamp recordings show blocking IKLV increased the excitability of the LES afferent terminals but not of S afferent terminals.
Furthermore, quantal and non-quantal modes of transmission are operational at the type I HC-calyx synapse, we characterize its precise encoding differences in utricular zones using the semi-intact utricle preparation (Songer & Eatock 2013) of CD-1 mice of ages P7 to P30, this allow us to patch-clamp the calyx terminal and record responses by stimulating the hair cell enveloped. The sine stimulus is applied via a stiff probe to study the quantal and non-quantal responses at different frequency stimulations to measure the vector strength and phase of the responses (fig. 2). Additionally, using sulforhodamine 101 or AlexaFluor 594 we capture the morphology of the afferent terminals recorded (fig. 3) in order to correlate terminal morphology with function.

Figure 2. Voltage responses of calyces to the sine stimuli (bottom) of the type I hair cell in S (A) and LES (B). The response has a larger non-quantal component in S/MES (16 mV p-p at 2 Hz) than in LES (4.8 mV p-p at 2 Hz). Mid frequencies (5, 10, 20 and 40 Hz) stimulus was sufficient to evoked spikes in S/MES, while in all frequencies evoked spikes in LES. C and D, phase histograms of spike responses from afferents in A and B, respectively, show high vector strength at mid frequencies for A and for B higher vector strength at higher frequencies (40, 50, 80, 100 Hz). E, bode plots of the calyx responses, the S/MES response it is highly tuned to mid frequencies, were it also maintained a phase lead. The LES response was broadly tuned from 10 to 100 Hz, with phase lead up to 50 Hz.
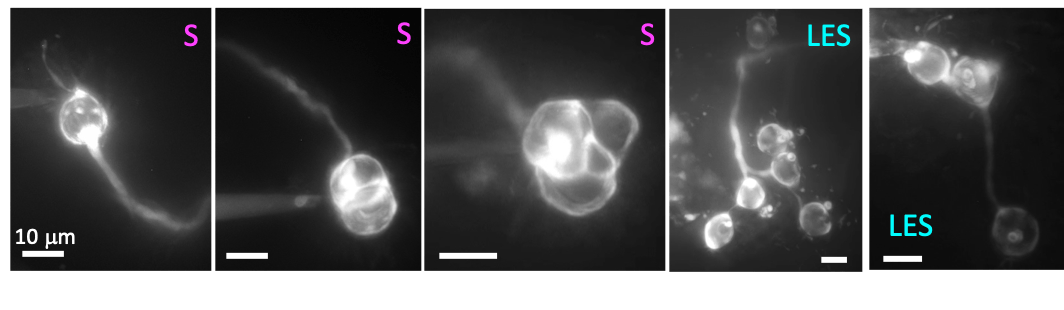
Figure 3. Diversity of afferent terminals in the utricle. Sulforhodamine filled afferent show different morphologies, calyx only afferents are present mainly in striolar region (S), while dimorphic afferent are present throughout the whole epithelia, some examples are shown from the lateral extrastriolar region (LES).
References
Contini D, Price SD, Art JJ (2017) Accumulation of K in the synaptic cleft modulates activity by influencing both vestibular hair cell and calyx afferent in the turtle. J of Physiol 595.3:777–803.
Goldberg JM, Desmadryl G, Baird RA, Fernández C (1990) The vestibular nerve of the chinchilla. V. Relation between afferent discharge properties and peripheral innervation patterns in the utricular macula. J Neurophysiol 63(4):791-804.
Horwitz GC, Risner-Janiczek JR, Holt JR (2014) Mechanotransduction and hyperpolarization-activated currents contribute to spontaneous activity in mouse vestibular ganglion neurons. J Gen Physiol 143(4):481-497.
Kalluri R, Xue J, Eatock RA (2010) Ion channels set spike timing regularity of mammalian vestibular afferent neurons. J Neurophysiol 104:2034-2050.
Lysakowski A, Gaboyard-Niay S, Calin-Jageman I, Chatlani S, Price SD, Eatock RA (2011) Molecular microdomains in a sensory terminal, the vestibular calyx ending. J Neurosci 31:10101-10114.
Meredith FL, Rennie KJ (2015) Zonal variations in K currents in vestibular crista calyx terminals. J Neurophysiol 113:264-276.
Songer, JE and Eatock RA (2013) Tuning and timing in mammalian type I hair cells and calyceal synapses. J Neurosci 33(8):3706-3724.
Pérez C, Limón A, Vega R, Soto E (2009) The muscarinic inhibition of the potassium M-current modulates the action-potential discharge in the vestibular primary-afferent neurons of the rat. Neuroscience 158:1662-1647.